Arq. Bras. Oftalmol. 2019; 82 (6): 10.5935/0004-2749.20190102
Total: 1577
Abdullah Ilhan1; Salih Altun2; Irfan Durukan1; Umit Yolcu1; Uzeyir Erdem2
DOI: 10.5935/0004-2749.20190102
ABSTRACT
PURPOSE: To investigate the potential associations between keratoconus and catalase rs1001179, superoxide dismutase 2 rs4880, and glutathione peroxidase 1 rs1050450 gene polymorphisms in a Turkish population.
METHODS: The study group included 121 unrelated keratoconus patients and 94 unrelated healthy controls. Blood samples (200 ml) were collected from all patients and controls to isolate genomic DNA. Genotyping was performed to identify rs1001179, rs4880, and rs1050450 using real-time polymerase chain reaction (PCR). Genotype and allele frequencies were calculated; their associations with keratoconus risk were assayed, and the association with keratoconus risk and demographic factors was examined.
RESULTS: Glutathione peroxidase 1 rs1050450 polymorphism was present in 41% cases compared with 29% controls (OR=1.66; 95% CI=1.11-2.50; p=0.014). No association was observed between catalase rs1001179 and SOD2 rs4880 polymorphisms and keratoconus (for all, p>0.05).
CONCLUSIONS: This study evaluated possible relationships between rs1050450, rs1001179, and rs4880 polymorphisms and keratoconus susceptibility. We found a possible association between glutathione peroxidase 1 rs1050450 polymorphism and an increased risk of keratoconus. However, the genotype and allele frequencies were identical in the catalase rs1001179 and superoxide dismutase 2 rs4880 polymorphisms. Further studies are needed to analyze the effect of such variations in identifying keratoconus susceptibility.
Keywords: Keratoconus; Glutathione peroxidase; Catalase; Superoxide dismutase; Polymorphism, genetic
RESUMO
OBJETIVO: Investigar as possíveis associações entre o ceratocone e os polimorfismos rs1001179 da catalase, rs4880 da superóxido-dismutase 2 e rs1050450 da glutationa-peroxidase 1 rs1050450 em uma população turca.
MÉTODOS: O grupo de estudo incluiu 121 pacientes com ceratocone não relacionados e 94 controles saudáveis também sem pa rentesco. Amostra de sangue (200 mL) foram coletadas de todos os pacientes e controle para isolar o DNA genômico. A genotipagem foi realizada para identificar rs1001179, rs4880 e rs1050450 utilizando a reação em cadeia da polimerase (PCR) em tempo real. As frequências de genótipos e alelos foram calculadas, suas associações com o risco de ceratocone foram avaliadas, e a associação com risco de ceratocone e fatores demográficos foi examinada.
RESULTADOS: O polimorfismo da glutationa-peroxidase 1 rs1050450 estava presente em 41% dos casos, comparado com 29% dos controles (OR=1,66, IC 95%=1,11-2,50; p=0,014). Não foi observada associação entre o ceratocone e os polimorfismos rs1001179 e SOD2 rs4880 da catalase (para todos, p>0,05).
CONCLUSÕES: Este estudo avaliou possíveis relações entre os polimorfismos rs1001179, rs4880 e suscetibilidade a cerato cone. Encontramos uma possível associação entre po limorfis mo da glutationa-peroxidase 1 rs1050450 e um risco aumentado de ceratocone. No entanto, o genótipo e as frequências alélicas foram idênticas nos polimorfismos rs1001179 da catalase e superóxido-dismutase 2 rs4880. Mais estudos são necessários para esclarecer o efeito dessas va riações na detecção da sus cetibilidade ao ceratocone.
Descritores: Ceratocone; Glutationa peroxidase; Catalase; Superóxido dismutase; polimorfismo genético
INTRODUCTION
Keratoconus (KC) is a progressive degenerative collagen disorder distinguished by stromal thinning, corneal steepening, and irregular astigmatism leading to deteriorated visual acuity and impaired quality of life. It mainly occurs in the second decade of life at an incidence of approximately 1 per 2000 in the general population. KC affects all ethnicities and both genders1.
The etiology of KC remains unclear. Various factors contribute to the pathophysiological processes of KC, including environmental, behavioral, and genetic factors. As 6% to 10% of KC patients have familial KC, genetic susceptibility has been widely studied for years. However, no specific genetic defect for KC has been described. In addition, several investigations have acknowledged the role of oxidative stress in KC pathogenesis. KC corneas accumulate cytotoxic end products of the nitric oxide and lipid peroxidation pathways and have impaired antioxidant enzymes and increased mitochondrial DNA damage2-7. These findings suggest that oxidative stress is significantly involved in the pathogenesis of KC. However, additional genetic and molecular studies are needed to thoroughly define the underlying problem.
Under physiological conditions of aerobic organisms, reactive oxygen species (ROS) generated during the course of metabolism is restricted by antioxidant enzymes. Superoxide, the primary ROS, is converted to hydrogen peroxide (H2O2), mainly by superoxide dismutase (SOD). H2O2 is decomposed by catalase (CAT) or glutathione peroxidase (GPX), releasing water and molecular oxygen. Any malfunction, i.e., in genetic polymorphisms, in these enzymes could lead to a shift in the balance between ROS production and the antioxidant defense mechanism resulting in oxidative stress8. SOD, CAT, and GPX have been described in aqueous humor, indicating their significance in the balance of oxidative status in the eyes9.
SOD has 3 subtypes. SOD1 and SOD2 are intracellular subtypes, and SOD3 is extracellular; all are present in the cornea4,10. SOD1 has been evaluated numerous times, and many studies have shown potential causative roles of SOD111,12. CAT and GPX are upregulated in KC corneas, emphasizing their importance in de creasing ROS13,14.
Recently, more attention has been given to the importance of genomic variations between individuals. Single nucleotide polymorphism (SNP) can be detected at certain positions in the genome. SNPs in the gene coding may affect metabolic pathways by altering the enzyme structure or substrate affinity and/or activity and impairing antioxidant capacity.
Many studies have revealed phenotype associations for SOD2 (Val16Ala, rs4880) including heart diseases, several malignancies, fatty liver disease, and age-related macular degeneration. Polymorphism studies of the CAT (-262C>T, rs1001179) gene have been conducted in many diseases including diabetes, systemic lupus erythematosus, Alzheimer disease, and several malignancies. The GPX-1 (Pro198Leu, rs1050450) polymorphism has also been extensively studied, with significant results being reported in panic disorder, kidney disease, morbid obesity, and cardiovascular diseases. However, no reports have described these common SNPs in KC patients. In fact, no study has yet reported on SOD2, GPX, and CAT SNPs in KC.
We aimed to determine whether the presence of these SNPs in key antioxidant enzymes is associated with an increased risk of KC.
METHODS
Patients and controls
Our study enrolled 121 unrelated Turkish patients with KC (aged 14 to 62 years) and 94 age-matched individuals with no history of KC as the control group. Each subject granted informed consent before study participation. The principles of the Declaration of Helsinki were strictly followed during all stages of the study.
All patients and controls were evaluated at the Department of Ophthalmology, Gulhane Military Medical Academy. The diagnosis of KC was made based on the clinical signs (Munson sign, protrusion, Vogt striae, corneal thickness, scarring, Fleischer rings, refractive errors) and topographic findings obtained using Oculus Pentacam (OCULUS Optikgeräte GmbH, Wetzlar, Germany) and OPD-Scan ARK-10000 (NIDEK, Tokyo, Japan).
The patients with a confirmed diagnosis of KC were included in the study group. Control subjects underwent the same examinations to eliminate mild cases that could be overlooked in regular examinations. None of the participants reported any genetic disorder.
GPX1 rs1050450, SOD2 rs4880, and CAT rs1001179 genotyping
Peripheral blood samples were collected in EDTA tubes and stored at -20°C before use. Genomic DNA was extracted from the blood samples (200 ml) using the EZ1 DNA Blood 200 ml Kit by EZ1 Advanced XL automated purification system (Qiagen, Hilden, Germany) according to the manufacturer’s instructions.
The SNP identification assay (LightSNiP; Tib-MolBiol, Berlin, Germany), which is based on simple probe melting curve analysis, was used to detect GPX1 (rs1050450), SOD2 (rs4880), and CAT (rs1001179) polymorphisms. Both PCR and melting curve analysis were conducted on the Rotor-Gene Q (Qiagen). The reaction for each SNP was performed in a final volume of 20 ml and included 2 ml DNA, 1 µl LightSNiP reagent mix (Tib-MolBiol), 2 ml LightCycler FastStart DNA Master HybProbe (Roche Diagnostics GmbH, Mannheim, Germany), 1.6 ml Mg Cl2 (25 mM), and 14.4 ml H2O. PCR consisted of initial denaturation at 95°C for 10 minutes came next by 45 cycles, consisting of denaturation at 95°C for 10 seconds, annealing at 60°C for 10 seconds, and elongation at 72°C for 15 seconds. After the last cycle, samples were held at 95°C for 30 seconds before the temperature was lowered to 45°C. Next, melting was conducted by steadily raising the temperature in 0.5°C increments every 2 seconds from 45°C to 75°C.
Statistical analysis
Hardy-Weinberg equilibrium was evaluated using the chi-squared (c2) test. The c2 test and two-way Student’s t-test were used to conduct statistical comparisons of genotypic and allelic distributions between the control and KC patients. All statistical analyses were performed using SPSS for Windows, version 16.0 (SPSS Inc., Chi cago, IL, USA). A p value £0.05 was considered statis tically significant.
RESULTS
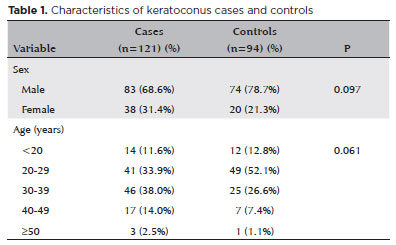
The frequency distribution of selected characteristics of the cases and controls is shown in table 1. No statistically significant difference was seen in sex or age distribution between the cases and control (both p>0.05).
Distributions of the GPX1 rs1050450 C/T, SOD2 rs4880 T/C, and CAT rs1001179 C/T genotypes consistent with the Hardy-Weinberg equilibrium among patients (p=0.867, 0.554, 0.199, respectively) and controls (p=0.901, 0.921, 0.839, respectively). We examined the genetic polymorphisms in the GPX1, SOD2, and CAT genes, and allele and genotype distributions are shown in table 2.
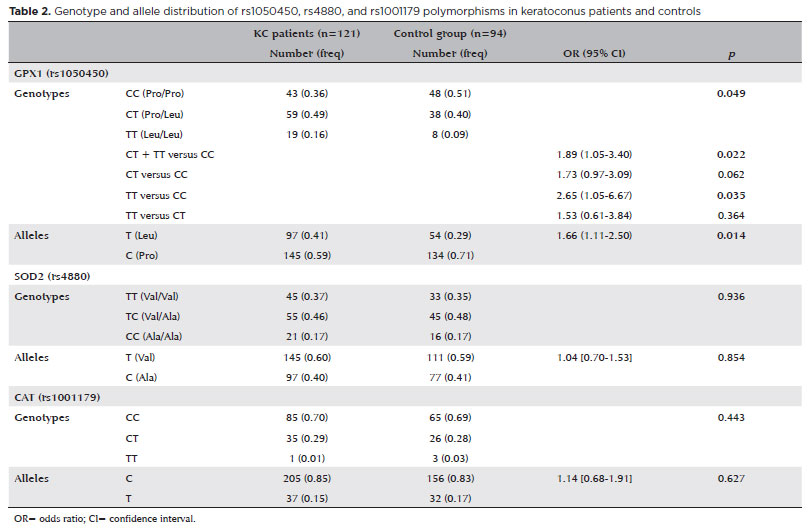
No significant difference existed between the SOD2 (rs4880) and CAT (rs1001179) genotypes (p=0.936 and 0.443, respectively) or allele frequencies (p=0.854 and 0.627, respectively) of the KC patients and the controls.
The difference in the allele frequency of the variant T allele of the GPX1 C/T polymorphism was statistically significant in the KC patients (41%) when compared with the controls (29%) (OR=1.66; 95% CI=1.11-2.50; p=0.014), thus the T allele was a statistically significant risk factor for KC. In KC patients, the GPX1 C/T polymorphism was present in 43 (36%) wild-type for the C/C genotype, 59 (49%) heterozygous for the C/T genotype, and 19 (16%) homozygous for the T/T genotypes. Statistical analysis demonstrated a possible harmful effect of the GPX1 C/T+TT and TT genotypes (OR=1.89, 95% CI=1.05-3.40; p=0.022; OR=2.65; 95% CI=1.05-6.67; p=0.035, respectively) in the development of KC and a possible protective effect of the C/C genotype in controls. Table 3 presents the comparative analysis of the demographic data and the SNPs.
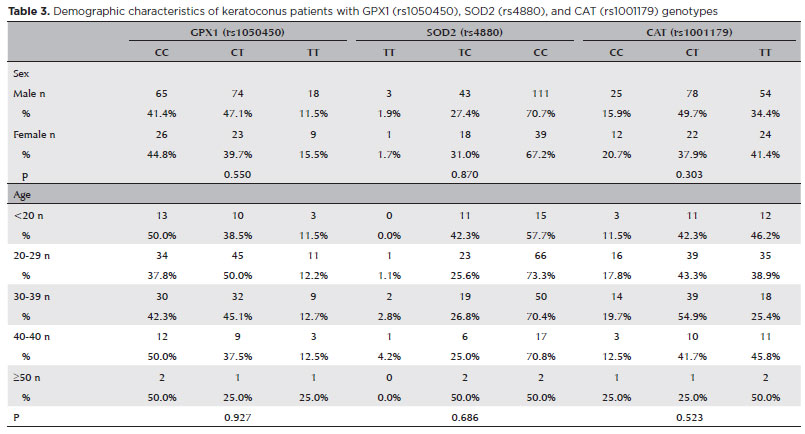
In our study, the GPX1 (rs1050450), SOD2 (rs4880), and CAT (rs1001179) polymorphisms did not impact the risk of KC with respect to sex (p=0.550, 0.870, 0.303, respectively) and age (p=0.927, 0.686, 0.523, respectively).
DISCUSSION
Our main concern in researching CAT rs1001179, SOD2 rs4880, and GPX1 rs1050450 polymorphisms in KC pathogenesis stems from their distinct enzymatic properties. They are essential to limiting ROS accumulation and inhibiting premature apoptosis in eukaryotic cells. In addition, they exist in almost all living organisms that require oxygen. SOD2, GPX1, and CAT share a common metabolic detoxification pathway15. Dysfunctions of GPX1, SOD2, and CAT are shown to play an integral role in the pathogenesis of various diseases. In addition, previous studies have reported oxidative stress and oxidative stress-induced premature apoptosis. Thus, an evaluation of these antioxidant enzymes would provide essential data5. Finally, after extensively searching the literature, we did not find a report on the role of SNPs associated with these enzymes in KC pathogenesis. These factors motivated us to evaluate the possible association of these SNPs with KC patients of Turkish origin. In the present study, we evaluated the role of rs1050450 (GPX1), rs4880 (SOD2), and rs1001179 (CAT) polymorphisms in detecting KC susceptibility. We chose to assess the most prominently researched polymorphisms that were compatible with multiple disorders16-18.
We identified a significant association between the rs1050450 polymorphism of GPX1 genotype and KC. GPX1 is involved in the decomposition H2O2 into water and oxygen. It is one of the most significant antioxidant enzymes in higher vertebrates and humans and is present in both cytoplasm and mitochondria. Brigelius-Flohé et al. reported that GPX1 could not compensate for other selenoproteins in the elimination of systemic oxidative stress, making it a primary antioxidant enzyme in vivo19. GPX1 also accelerates the reduction of lipid peroxides and hydrogen as well as peroxynitrite20. Therefore, GPX1 dysfunction may cause the accumulation of peroxynitrite and lipid hydroperoxides. The extreme amount of reactive oxygen and nitric species may damage nucleic acids, membrane lipids, mitochondrial DNA, and proteins. Various studies have reported that this metabolite accumulation may induce apoptosis21,22. Further, keratocyte apoptosis is seen in keratoconic corneas23,24. Based on these facts and our results, we propose that GPX1 dysfunctions should be considered in KC pathogenesis.
No statistically significant association existed between the rs1001179 polymorphism of the CAT gene, rs4880 polymorphism of the SOD2 gene, and KC risk. Many studies on different diseases have reported both significant and insignificant results for both SNPs25-28. The genotype distribution of these SNPs fluctuates between populations29,30. These discrepancies are probably due to the limited number of subjects and population and/or ethnicity differences.
As the genotype and allele frequency of both polymorphisms were identical, it is assumed that the rs1001179 polymorphism of the CAT gene and rs4880 polymorphism of the SOD2 gene are not involved in KC pathogenesis. Our study was conducted in the Turkish population, and similar studies are needed in different populations to precisely define the relationship between these SNPs and KC.
To the best of our knowledge, this is the first report to investigate the potential associations between the rs1050450, rs1001179, and rs4880 polymorphisms and KC susceptibility. In conclusion, we confirmed a possible association of the GPX1 rs1050450 polymorphism with KC in the Turkish population. However, we did not observe a significant difference in the CAT rs1001179 and SOD2 rs4880 polymorphisms between the study and control groups. Further studies with large cohorts of multiple nationalities and ethnicities are needed to clarify the association and significance of this metabolic pathway.
REFERENCES
1. Gordon-Shaag A, Millodot M, Shneor E, Liu Y. The genetic and environmental factors for keratoconus. BioMed Res Int. 2015;2015:795738.
2. Buddi R, Lin B, Atilano SR, Zorapapel NC, Kenney MC, Brown DJ. Evidence of oxidative stress in human corneal diseases. J Histochem Cytochem. 2002;50(3):341-51.
3. Gondhowiardjo TD, van Haeringen NJ, Völker-Dieben HJ, Beekhuis HW, Kok JH, van Rij G, et al. Analysis of corneal aldehyde dehydrogenase patterns in pathologic corneas. Cornea. 1993;12(2):146-54.
4. Behndig A, Karlsson K, Johansson BO, Brännström T, Marklund SL. Superoxide dismutase isoenzymes in the normal and diseased human cornea. Invest Ophthalmol Vis Sci. 2001;42(10):2293-6.
5. Kenney MC, Chwa M, Atilano SR, Tran A, Carballo M, Saghizadeh M, et al. Increased levels of catalase and cathepsin V/L2 but decreased TIMP-1 in keratoconus corneas: evidence that oxidative stress plays a role in this disorder. Invest Ophthalmol Vis Sci. 2005;46(3):823-32.
6. Atilano SR, Coskun P, Chwa M, Jordan N, Reddy V, Le K, et al. Accu¬mulation of mitochondrial DNA damage in keratoconus corneas. Invest Ophthalmol Vis Sci. 2005;46(4):1256-63.
7. Göncü T, Akal A, Adibelli FM, Çakmak S, Sezen H, Yilmaz OF. Tear Film and Serum Prolidase Activity and Oxidative Stress in Patients With Keratoconus. Cornea. 2015;34(9):1019-23.
8. Lubrano V, Balzan S. Enzymatic antioxidant system in vascular inflammation and coronary artery disease. World J Exp Med. 2015; 5(4):218-24.
9. Abu-Amero KK, Kondkar AA, Mousa A, Osman EA, Al-Obeidan SA. Analysis of catalase SNP rs1001179 in Saudi patients with primary open angle glaucoma. Ophthalmic Genet. 2013;34(4):223-8.
10. Liu C, Ogando D, Bonanno JA. SOD2 contributes to anti-oxidative capacity in rabbit corneal endothelial cells. Mol Vis. 2011;17: 2473-81.
11. Moschos MM, Kokolakis N, Gazouli M, Chatziralli IP, Droutsas D, Anagnou NP, et al. Polymorphism Analysis of VSX1 and SOD1 Genes in Greek Patients with Keratoconus. Ophthalmic Genet. 2015;36(3):213-7.
12. Udar N, Atilano SR, Brown DJ, Holguin B, Small K, Nesburn AB, et al. SOD1: a candidate gene for keratoconus. Invest Ophthalmol Vis Sci. 2006;47(8):3345-51.
13. Chwa M, Atilano SR, Reddy V, Jordan N, Kim DW, Kenney MC. Increased stress-induced generation of reactive oxygen species and apoptosis in human keratoconus fibroblasts. Invest Ophthalmol Vis Sci. 2006;47(5):1902-10.
14. Leema G, Muralidharan AR, Annadurai T, Kaliamurthy J, Geraldine P, Thomas PA. Oxidative stress in experimental rodent corneas infected with aflatoxigenic and nonaflatoxigenic Aspergillus flavus. Cornea. 2013;32(6):867-74.
15. Bastaki M, Huen K, Manzanillo P, Chande N, Chen C, Balmes JR, et al. Genotype-activity relationship for Mn-superoxide dismutase, glutathione peroxidase 1 and catalase in humans. Pharmacogenet Genomics. 2006;16(4):279-86.
16. Sousa VC, Carmo RF, Vasconcelos LR, Aroucha DC, Pereira LM, Moura P, et al. Association of Catalase and Glutathione Peroxidase 1 Polymorphisms with Chronic Hepatitis C Outcome. Ann Hum Genet. 2016;80(3):145-53.
17. Kidir V, Uz E, Yigit A, Altuntas A, Yigit B, Inal S, et al. Manganese superoxide dismutase, glutathione peroxidase and catalase gene polymorphisms and clinical outcomes in acute kidney injury. Ren Fail. 2016;38(3):372-7.
18. Kakkoura MG, Demetriou CA, Loizidou MA, Loucaides G, Neophytou I, Malas S,et al. MnSOD and CAT polymorphisms modulate the effect of the Mediterranean diet on breast cancer risk among Greek-Cypriot women. Eur J Nutr. 2016;55(4):1535-44.
19. Brigelius-Flohé R, Maiorino M. Glutathione peroxidases. Biochim Biophys Acta. 2013;1830(5):3289-303. Review.
20. Tan SM, Stefanovic N, Tan G, Wilkinson-Berka JL, de Haan JB. Lack of the antioxidant glutathione peroxidase-1 (GPx1) exacerbates retinopathy of prematurity in mice. Invest Ophthalmol Vis Sci. 2013;54(1):555-62.
21. Yang WS, Stockwell BR. Ferroptosis: Death by Lipid Peroxidation. Trends Cell Biol. 2016;26(3):165-76.
22. Das UN. Essential fatty acids, lipid peroxidation and apoptosis. Prostaglandins Leukot Essent Fatty Acids. 1999;61(3):157-63.
23. Palamar M, Onay H, Ozdemir TR, Arslan E, Egrilmez S, Ozkinay F, et al. Relationship between IL1ß-511C>T and ILRN VNTR polymorphisms and keratoconus. Cornea. 2014;33(2):145-7.
24. Wójcik KA, Blasiak J, Kurowska AK, Szaflik J, Szaflik JP. [Oxidative stress in the pathogenesis of keratoconus]. Klin Oczna. 2013; 115(4):311-6. Review. Polish.
25. Rajic V, Aplenc R, Debeljak M, Prestor VV, Karas-Kuzelicki N, Mlinaric-Rascan I, et al. Influence of the polymorphism in candidate genes on late cardiac damage in patients treated due to acute leukemia in childhood. Leuk Lymphoma. 2009;50(10):1693-8.
26. Góth L, Nagy T, Kósa Z, Fejes Z, Bhattoa HP, Paragh G, et al. Effects of rs769217 and rs1001179 polymorphisms of catalase gene on blood catalase, carbohydrate and lipid biomarkers in diabetes mellitus. Free Radic Res. 2012;46(10):1249-57.
27. Kondo N, Bessho H, Honda S, Negi A. SOD2 gene polymorphisms in neovascular age-related macular degeneration and polypoidal choroidal vasculopathy. Mol Vis. 2009;15:1819-26.
28. Méplan C, Hughes DJ, Pardini B, Naccarati A, Soucek P, Vodickova L, et al. Genetic variants in selenoprotein genes increase risk of colorectal cancer. Carcinogenesis. 2010;31(6):1074-9.
29. Silig Y, Tas A. Manganese-superoxide dismutase (MnSOD) polymorphisms. Turk J Biochem. 2015;40(2):163-8.
30. Sutton A, Khoury H, Prip-Buus C, Cepanec C, Pessayre D, Degoul F. The Ala16Val genetic dimorphism modulates the import of human manganese superoxide dismutase into rat liver mitochondria. Pharmacogenetics. 2003;13(3):145-57.
Submitted for publication:
March 20, 2017.
Accepted for publication:
May 2, 2019.
Approved by the following research ethics committee: Ataturk University Faculty of Medicine (#B.30.2.ATA.0.01.00/73).
Funding: No specific financial support was available for this study.
Disclosure of potential conflicts of interest: None of the authors have any potential conflicts of interest to disclose.

How to cite this article:
 ABO is licensed under a Creative Commons Attribution-NonComercial 4.0 Internacional.
ABO is licensed under a Creative Commons Attribution-NonComercial 4.0 Internacional.