Arq. Bras. Oftalmol. 2004; 67 (1): 10.1590/S0004-27492004000100032
Total: 4579
Harley E. A. Bicas1
DOI: 10.1590/S0004-27492004000100032
RESUMO
A visão binocular de seres humanos resulta da superposição quase completa dos campos visuais de cada olho, o que suscita discriminação perceptual de localizações espaciais de objetos relativamente ao observador (localização egocêntrica) bem mais fina (estereopsia), mas isso ocorre em, apenas, uma faixa muito estreita (o horóptero). Aquém e além dela, acham-se presentes diplopia e confusão, sendo necessária supressão fisiológica (cortical) para evitá-las. Analisa-se a geometria do horóptero e suas implicações fisiológicas (o desvio de Hillebrand, a partição de Kundt, a área de Panum), assim como aspectos clínicos da visão binocular normal (percepção simultânea, fusão, visão estereoscópica) e de adaptações a seus estados afetados (supressão patológica, ambliopia, correspondência visual anômala).
Descritores: Visão binocular; Ambliopia; Transtornos da visão; Percepção visual; Percepção de profundidade
ABSTRACT
The binocular vision of human beings is given by the almost complete superimposition of the monocular visual fields, which allows a finer perceptual discrimination of the egocentric localization of objects in space (stereopsis) but only within a very narrow band (the horopter). Before and beyond it, diplopia and confusion are present, so that a physiologic (cortical) suppression is necessary to avoid them to become conscious. The geometry of the horopter and its physiologic implications (Hillebrand's deviation, Kundt's partition, Panum's area, stereoscopic vision) are analyzed, as well as some clinical aspects of the normal binocular vision (simultaneous perception, fusion, stereoscopic vision) and of adaptations to abnormal states (pathologic suppression, amblyopia, abnormal retinal correspondence).
Keywords: Vision, binocular; Amblyopia; Vision disorders; Visual perception; Depth perception
INTRODUÇÃO
Binocularidade, em seu sentido mais amplo, é o termo que se aplica à capacitação de apreender estímulos visuais com dois olhos. Entre as diferentes espécies que possuem esse atributo, os modelos da relação binocular, suas peculiaridades e fins são, entretanto, muito distintos. Enquanto para coelhos a função binocular é a de provimento de campos visuais independentes, garantidos por órgãos visuais de cada lado da cabeça, nos primatas superiores (e no homem), a percepção visual do espaço se dá com base na frontalização dos olhos. Assim, coelhos gozam de uma extensa visão de "campo", cobrindo 360º, um escrutínio completo do espaço ao redor de seu corpo(1), o que lhes propicia meios de fuga de predadores(2). Já entre nós predomina a superposição praticamente completa dos campos visuais. Tal superposição, ocasionando perda de 180º da discriminação visual do espaço (relativamente à do coelho) traria como vantagem, em contrapartida, a capacidade de percepção de "profundidades" de objetos nesse campo visual, isto é, a de percepção das localizações egocêntricas desses objetos (à distância deles ao agente da percepção).
Tal assertiva de "vantagem filogenética" identifica-se como viés antropocêntrico e pode ser tomada como argumento equivocado a que faltam sustentações, tanto de causas como de efeitos. De fato, a percepção de distâncias egocêntricas não é necessariamente vinculada a essa superposição de campos, podendo ser dada por uma série de "pistas monoculares" tais como as de:
a) interposição de estímulos (os mais próximos "cobrindo" os contornos e áreas dos mais distantes);
b) tamanhos relativos das imagens (maiores para os de objetos mais próximos, menores para os dos mais distantes);
c) contornos e brilhos (mais acentuados com a proximidade, esmaecidos com o distanciamento);
d) zonas de sombras e iluminação (sugerindo relevos e cavidades);
e) perspectiva aérea (coloração mais azulada para grandes distâncias, pela interposição de ar entre o observador e os objetos);
f) perspectiva cinemática (pelo observador em movimento: objetos mais próximos com deslocamentos aparentes mais rápidos).
Ademais, mecanismos acomodativos especializados poderiam dar conta dessa discriminação de distâncias egocêntricas.
Por outro lado, a superposição de campos não assegura garantia de discriminação do distanciamento egocêntrico de todos os elementos do espaço percebido. Ao contrário, como será visto, essa propriedade é relacionada, apenas, a uma estreita zona, cabendo mecanismos de supressão da binocularidade para as demais(1), uma "inibição facultativa" (termo criado por Worth)(3). Ou seja, a superposição de campos visuais, tratada como modelo para obtenção de benefícios perceptuais acaba exigindo, de modo paradoxal, que as informações não sejam simultânea e ubiqüitariamente aproveitadas.
De qualquer modo, é esse tipo de visão binocular, com suas particularidades e condicionamentos que nos cabe privilegiar por tratamentos.
A) Percepção monocular do espaço
Elementos materiais do espaço objetivo emitem ou refletem luz, que, atravessando as estruturas oculares, forma-lhes as imagens ópticas na retina. O verdadeiro trajeto das frentes de onda da energia eletromagnética é, então, composto por um complicado conjunto de linhas, com direções mudadas a cada refração ocorrida nas variadas interfaces (dioptros) dos diferentes meios oculares de transmissão da luz. Mas uma linha única e abstrata pode ser traçada entre o considerado ponto objeto do espaço e sua respectiva imagem na retina(*).
A essa linha abstrata, mas unívoca, entre o objeto no espaço e sua imagem na retina, corresponde uma localização subjetiva do elemento gerador do estímulo, sua direção visual. Objetos cujas imagens formam-se sobre a fóvea, são percebidos "em frente" do olho, têm a chamada direção visual principal, ou linha visual principal (que tem, como correlato físico, o eixo visual), uma espécie de eixo referencial, relativamente ao qual as demais localizações espaciais são referidas. Objetos no campo visual nasal (N, figura 1a), têm suas imagens formadas na retina temporal (t , figura 1a), cuja percepção é a do objeto situado no respectivo lado nasal do olho (N¢, figura 1b). Objetos no campo superior (S, figura 1a), formam imagens na retina inferior (i, figura 1a) sendo a imagem então percebida do lado superior desse olho, etc (figura 1).
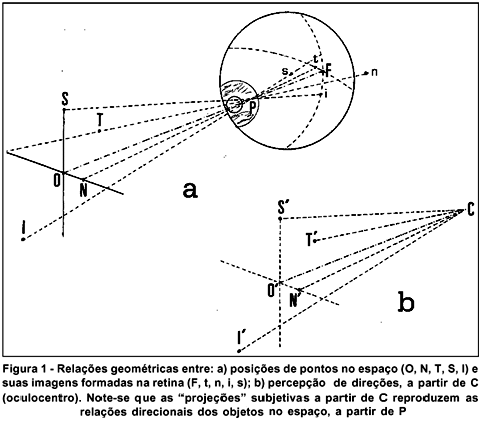
O ponto em que se cruzam as linhas de correlação espacial entre objetos e suas imagens (P, na figura 1a) é o ponto nodal do sistema óptico ocular(**). As relações angulares entre as posições espaciais dos objetos (O, N, T, S, I) ou de suas respectivas imagens (F, t, n, i, s) a partir do ponto nodal (P) são mantidas na reprodução perceptual das direções visuais (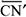 ,
, 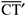 , etc), relativamente à principal(
, etc), relativamente à principal(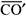 ) (Figura 1b). Em outras palavras, os ângulos visuais são medidos a partir dos pontos nodais(4). Na verdade, o centro de um círculo de difusão na retina é determinado pela linha pupilar (que passa pelo centro da pupila de saída e o foco posterior) e não pela linha que une o ponto nodal posterior com o foco posterior (linha visual)(4). Esse deve ter sido o motivo pelo qual um profundo conhecedor da fisiologia e da óptica ocular distingue localizações retínicas "relativas" e "absolutas", separa um "centro de divisão" e um "de projeção" das imagens e toma como referencial para o "centro de perspectiva" da retina, o centro da pupila de entrada do olho (cerca de 0,56 mm à frente do plano da íris, a 3,04 mm à frente do ponto nodal médio, que ficaria a 7,4 mm atrás da córnea)(5).
) (Figura 1b). Em outras palavras, os ângulos visuais são medidos a partir dos pontos nodais(4). Na verdade, o centro de um círculo de difusão na retina é determinado pela linha pupilar (que passa pelo centro da pupila de saída e o foco posterior) e não pela linha que une o ponto nodal posterior com o foco posterior (linha visual)(4). Esse deve ter sido o motivo pelo qual um profundo conhecedor da fisiologia e da óptica ocular distingue localizações retínicas "relativas" e "absolutas", separa um "centro de divisão" e um "de projeção" das imagens e toma como referencial para o "centro de perspectiva" da retina, o centro da pupila de entrada do olho (cerca de 0,56 mm à frente do plano da íris, a 3,04 mm à frente do ponto nodal médio, que ficaria a 7,4 mm atrás da córnea)(5).
Ao ponto C dá-se o nome de oculocentro, uma concepção também abstrata, cuja posição pode ser equiparada à do ponto nodal do sistema óptico (não confundir com o centro geométrico do olho).
B) Percepção binocular do espaço
A visada de um ponto objeto no espaço é normalmente provida pelos dois olhos, simultaneamente, de tal sorte que suas imagens se formem sobre as fóveas (tendo, por conseguinte, as mesmas direções visuais "em frente"). Outros objetos poderão ter suas imagens formadas em pontos retínicos correspondentes, isto é, os com idênticas direções visuais (relativamente à principal), ou não. No primeiro caso, cada objeto (O, A, B, figura 2) será percebido de modo único, enquanto os que têm imagens formadas em locais retínicos não correspondentes (L, Q, figura 3a), serão notados em duas localizações espaciais (diplopia). Os objetos cujas imagens são formadas em locais retínicos correspondentes dispõem-se sobre uma linha imaginária, o círculo de Vieth e Müller, cujo traçado passa pelo ponto principal de fixação (O, figura 2) e pelos pontos nodais de cada olho (PE e PD). Tal círculo é a representação teórica do conceito de horóptero, a região do espaço cujos objetos nela situados estimulam pontos retínicos correspondentes, sendo então percebidos univocamente. Fora do horóptero (figura 3a), haverá diplopia (figura 3c). Para objetos situados além do horóptero (L, figura 3a), a diplopia é homônima: a imagem mais à direita (LD) é a formada pelo olho direito e a mais à esquerda (LE) formada pelo olho esquerdo (figura 3c)(***). Para objetos situados aquém do horóptero (e.g., o ponto Q, figura 3a), a diplopia é cruzada: a imagem mais à esquerda é a do olho direito (QD)e vice-versa (figura 3c).
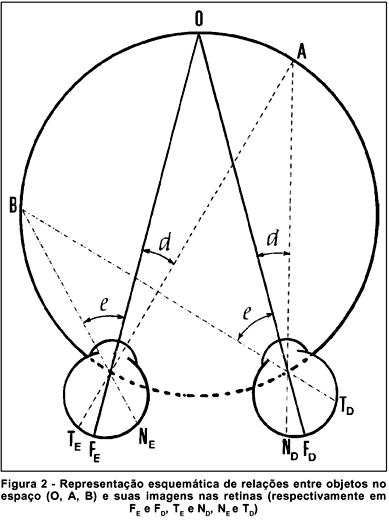
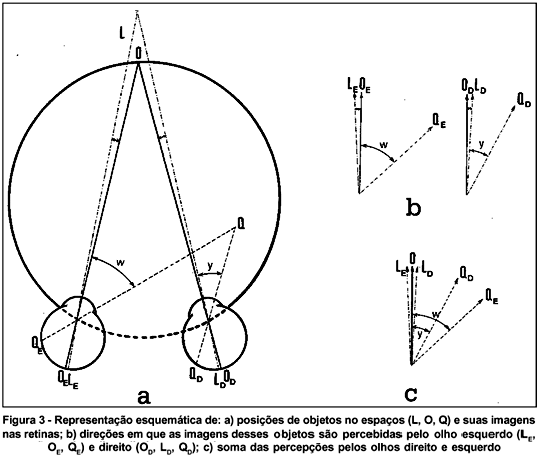
C) O horóptero real. Desvio de Hillebrand
Na verdade, o horóptero experimentalmente determinado difere do teórico. Costuma ser convexo (relativamente ao examinado) quando o ponto de fixação está a distâncias maiores do que (aproximadamente) dois metros; plano, para essa distância; e côncavo, para distâncias menores. Essa discrepância entre o horóptero teórico (sempre côncavo) e o obtido na prática é a do chamado desvio de Hillebrand e tem como explicação o fato de os pontos correspondentes não apresentarem disposição absolutamente simétrica com referência às respectivas fóveas(6) (figura 4).
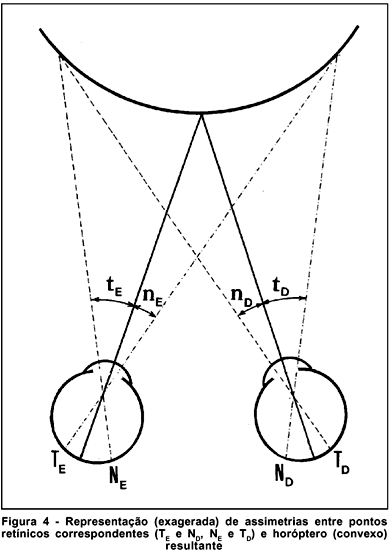
Daí resulta que, monocularmente, a divisão de uma linha ao meio não é feita de modo a que as duas metades fiquem idênticas. Em geral, a parte temporal da linha (correspondente à retina nasal) fica maior que a nasal (partição de Kundt). Discrepâncias até maiores são notadas na tentativa de divisão eqüitativa de uma linha vertical. Todavia, ainda não foi conclusivamente provado que a disposição dos campos receptivos correspondentes seja realmente (anatomicamente) assimétrica em relação às fóveas, havendo teorias que especulam com bases cognitivas para explicar esses fenômenos de partição e as anomalias da forma do horóptero(7).
D) Área de Panum
A concepção geométrica pela qual pontos do espaço são relacionados a pontos da retina, gerando linhas de correspondência visual (os horópteros) não é sustentável: os elementos retínicos responsáveis pela transdução dos estímulos físicos (luz) a sensações (o sinal neural) são os fotorreceptores e suas conexões, formando áreas relativamente extensas, os chamados campos receptivos. Mesmo na fóvea, em que o tamanho desses campos receptivos é mínimo (a área de secção transversal de cada cone foveal) a relação está, portanto, longe de ser puntiforme. Resulta que a cada um desses elementos fotorreceptores não cabe uma linha de direção visual, mas um feixe delas, formando uma figura cônica (figura 5a). Ao redor dos pontos que formariam a linha do horóptero (relacionada ao ponto central de cada um desses elementos), o entrecruzamento de tais feixes cônicos dos respectivos elementos de cada olho, perceptualmente correspondentes, gera um sólido, cuja secção no plano de estudo considerado (por exemplo, o horizontal) é uma superfície plana, a área de Panum (figura 5a). Pelo fato de os campos receptivos serem maiores na periferia do que na região central, a área de Panum não é uniforme. No centro é pequena (estimada em cerca de 15 minutos de arco, desde o limite interno ao externo)(8-9), estendendo-se à medida que regiões mais periféricas forem consideradas (figura 5b). Mas de qualquer modo, valores maiores para a área de Panum foram relatados também para a região central, na dependência do método usado(10). Os pontos do espaço contidos numa área de Panum delimitada pelas linhas de direção visual referentes aos contornos dos campos receptivos (elementos visuais) correspondentes possuem, pois, uma só, idêntica, direção visual. São então vistos unificadamente; e apenas além e aquém dessa superfície (e não de uma linha divisória do espaço) é que se tornaria demonstrável a diplopia.
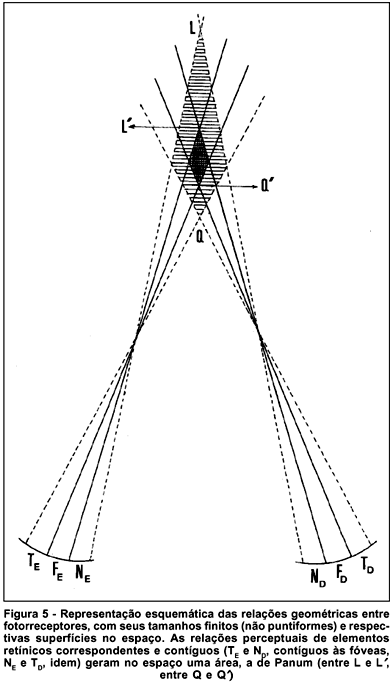
E) Estereopsia
Na verdade, há ainda outras zonas adicionais, contíguas à de Panum (à sua frente e atrás) nas quais a percepção diplópica, que deveria ser então demonstrada, não ocorre. É nessas "faixas adicionais" em que se origina o fenômeno da percepção estereoscópica, a de relevos e cavidades. Explica-se que quando o ângulo de separação das imagens de cada olho é muito reduzido, a diplopia resolve-se corticalmente como percepção estereoscópica: para a que seria homônima (além do horóptero) a noção é a de escavação, distanciamento (relativamente ao ponto de fixação) enquanto para a que seria cruzada (aquém do horóptero) surge a representação consciente de proximidade, relevo. Em outras palavras, elementos retínicos não correspondentes (mas contíguos aos verdadeiramente correspondentes, isto é, com idênticas direções visuais), também se relacionam corticalmente para gerar a percepção do espaço numa figuração estereoscópica.
Quanto menos discrepantes forem os distanciamentos angulares desses elementos não correspondentes, mais delicada é a sensação de cavidade ou ressalto e vice-versa. Estereopsias de 20 a 40² representam limites normais dessa percepção, mas separações angulares entre dois pontos de até cerca de 3000² (= 50¢) podem ainda ser ajuizadas como a de um único, com aproximação ou distanciamento.
F) Graus de binocularidade
Classicamente, admite-se que as funções binoculares podem ser graduadas segundo a maior ou menor hierarquia de seus desempenhos. Assim, o "primeiro" grau é o da percepção simultânea a estímulos (binoculares) das fóveas, ou das máculas, ou de regiões periféricas das retinas.
1) Percepção simultânea
Estritamente considerando, a decodificação de sinais emitidos por cada um dos olhos e seus processamentos simultâneos não são abonadas pelas teorias vigentes com as quais se entendem os mecanismos de integração cortical das sensações visuais (11). Por elas, a percepção de uma figura é admitida como resultando da composição espacial de seus pontos, uns devidos a estímulos ao olho direito, outros ao esquerdo (como no caso de pontos brancos e pretos formando um clichê), a teoria do mosaico; alternando-se temporalmente, enquanto varia a composição de cada contribuição monocular, a teoria da rivalidade retínica. Desse modo, seja por alternância espacial e, ou temporal, nunca o mesmo ponto objeto teria as suas imagens em cada um dos olhos simultaneamente processadas pelo sistema nervoso central. Contudo, está bem documentada a interação de campos receptivos binoculares num mesmo neurônio cortical, com mecanismos complexos de excitação e inibição. Por outro lado, "percepção" é o resultado subjetivo de tais mecanismos funcionais e a noção dela advinda é a de simultaneidade no funcionamento binocular. Logo, embora possa até inexistir uma "perfeita" simultaneidade perceptual binocular do ponto de vista fisiológico, o termo é cabível em sua interpretação psicofísica.
Na prática, o teste de percepção simultânea requer a apresentação independente de figuras distintas a cada olho. Aparelhos como o sinoptóforo prestam-se muito bem a essa demonstração: assim, a percepção binocular da figura de um soldado apresentada a um dos olhos e a de uma guarita ao outro, é relatada como a de um soldado "dentro" da guarita (enquadrado por ela). Em casos de supressão (ou quando o estado de rivalidade retínica mostra a dominância muito intensa de um dos olhos) apenas uma das imagens é percebida, eventualmente podendo haver alternância das mensagens mandadas ao córtex visual por cada olho, mas nunca sendo referida uma percepção simultânea.
Sem esse instrumento, a percepção simultânea pode ser comprovada, por exemplo, pelo relato de diplopia (espontânea, no estrabismo recente de um adulto, ou induzida pela anteposição de prismas aos olhos de um paciente normal), pelo teste com o vidro de Maddox (percepção simultânea de uma fonte luminosa por um olho e da imagem daquela, produzida pelo vidro canelado, diante do outro), pelos vidros de Bagolini, etc.
2) Fusão
Daí resulta que, monocularmente, a divisão de uma linha ao meio não é feita de modo a que as duas metades fiquem idênticas. Em geral, a parte temporal da linha (correspondente à retina nasal) fica maior que a nasal (partição de Kundt). Discrepâncias até maiores são notadas na tentativa de divisão eqüitativa de uma linha vertical. Todavia, ainda não foi conclusivamente provado que a disposição dos campos receptivos correspondentes seja realmente (anatomicamente) assimétrica em relação às fóveas, havendo teorias que especulam com bases cognitivas para explicar esses fenômenos de partição e as anomalias da forma do horóptero(7).
Uma vez demonstrada a capacidade de funcionamento binocular "simultâneo", pode-se examinar a de ajustamento a condições em que os estímulos não são dados precisamente a elementos correspondentes. Diz-se, então, testar a capacidade de fusão e a da amplitude em que ela pode ser demonstrada.
No sinoptóforo, as figuras usadas para testar fusão são as que se apresentam essencialmente idênticas, variando apenas em pequenas minúcias para cada olho. Por exemplo: para o direito a figura de um menino, segurando um ramalhete de flores com a mão direita, tendo a esquerda sem nada; para o olho esquerdo o mesmo menino em todos os detalhes, mas sustentando a alça de um balde com a mão esquerda, enquanto a direita nada porta. Com tal condição, a percepção binocular é a de superposição das figuras do menino, de modo que ele seja referido como único, segurando um ramalhete de flores e, ao mesmo tempo, um balde, com suas mãos direita e esquerda, respectivamente. Como preponderam os elementos comuns da imagem cabente a cada olho, há uma fusão deles; de modo que mesmo quando se procura deslocar um dos estímulos relativamente ao outro, ou ambos ao mesmo tempo, os olhos também se movem, "ajustam-se" ao deslocamento produzido das imagens retínicas, para mantê-las centradas nas respectivas fóveas. Diz-se ocorrer um movimento (binocular) fusional, disjuntivo (vergencial), cuja amplitude pode ser registrada. Se a tentativa de avaliação dessa função visual for feita com figuras que não a suscitam (por exemplo, a do soldado e a da guarita em que, embora possa ser demonstrada a percepção simultânea delas, não há componentes comuns que as conciliem numa "unidade"), não se consegue a resposta fusional vergencial. Claro, também, que se o deslocamento relativo das imagens estiver acima de certa velocidade (ao redor de 20 º/s), a vergência fusional também não ocorre.
O estudo da fusão pode ser feito com a fixação binocular a um ponto luminoso ou a objetos de uma sala, enquanto se coloca um prisma diante de um dos olhos. A imagem do objeto por ele fixado será então deslocada da fóvea, produzindo diplopia, mas desde que haja capacitação fusional e o distanciamento das imagens não for muito grande, elas poderão ser novamente juntadas. Em seqüência, prismas de valores maiores podem então ser testados, até o limite em que a diplopia então suscitada não puder ser mais compensada.
A amplitude da fusão depende, aliás, de vários fatores, tais como o da natureza do estímulo (um ponto luminoso num quarto escuro é um estímulo fusional fraco, quando comparado ao da imagem de um aparelho de televisão, em uma sala com outros objetos, em que a riqueza de elementos fusionais centrais e periféricos das retinas justifica uma resposta binocular mais elaborada), o método utilizado (geralmente o sinoptóforo mostra amplitudes fusionais maiores que as obtidas com prismas, no espaço), a técnica (a transição seqüencial e rápida entre prismas tende a produzir respostas fusionais menores), etc. Obviamente, além da capacitação sensorial, é também necessária a disponibilidade oculomotora para a execução dos movimentos. Olhos cujos movimentos estiverem restringidos (por causas mecânicas, como as de contenção; ou funcionais, como as de uma paralisia muscular) terão reduzidas amplitudes fusionais, ainda que a capacidade sensorial de fazê-las possa ser normal.
A convergência fusional (medida com prismas de base temporal) é a mais desenvolvida. Valores de 30D ou mais são comumente demonstráveis. Em contrapartida, a divergência fusional é bem menor (cerca de 8D, com prismas de base nasal), enquanto as divergências verticais (prisma de base inferior no OD e, ou superior no OE; ou vice-versa; no primeiro caso para medir uma divergência vertical D/E; no segundo para uma divergência vertical E/D) são ainda normalmente mais reduzidas. Valores maiores, tanto para divergência horizontal, quanto para a vertical, são, entretanto encontrados em alguns desvios oculomotores (compensados, aparecendo como heteroforias).
3) Estereopsia
O "terceiro grau" da binocularidade é o da estereopsia, avaliada com os chamados estereogramas. Estes são, basicamente, figuras de um mesmo conjunto, tomadas sob diferentes ângulos ("pontos de vista") de modo que quando ambas são comparadas, nota-se que os elementos de cada uma possuem suas posições relativas discretamente dissimilares. São essas discrepâncias as que suscitam a percepção estereoscópica (do grego, "estereós", sólido, tridimensional). Boa parte dos testes é então baseada no princípio de que as imagens vistas por um olho (por exemplo, com a anteposição de um filtro vermelho ou de um polaróide numa certa orientação) não é vista pelo outro (com a anteposição de um filtro verde, ou de um polaróide na orientação perpendicular à anterior) e vice-versa.
Desenhos mais sofisticados, de modo a possibilitar uma fusão "espontânea" de elementos visuais com pequenas disparidades para cada olho, são também disponibilizados para testes.
Um modo relativamente grosseiro, mas muito simples, de se examinar a capacidade de discriminação de distâncias egocêntricas é a de resposta do examinado à sua tentativa de fazer coincidir a ponta de uma haste (ou caneta, ou dedo) por ele dirigida, à ponta de outra (ou caneta, ou dedo) sustentada pelo examinador, no espaço à sua frente.
G) Adaptações sensoriais ao estado do desvio oculomotor
1) Supressões
Nos estrabismos, o direcionamento inadequado dos eixos visuais ao objeto de atenção é nítida; faz surgir a percepção dele em diferentes situações do espaço, a diplopia. Uma das imagens, entretanto, a situada sobre a fóvea do olho que fixa diretamente o objeto de atenção é nítida; enquanto a do olho desviado (relativamente a esse objeto) oferece formas menos discrimináveis (já que a imagem é originada de área retínica com menor acuidade visual). Mas, também, esse olho (desviado relativamente ao objeto de atenção) tem sua fóvea direcionada a outro ponto do espaço, cuja imagem será então percebida em frente. E daí, em visão binocular, notada como superposta à imagem do objeto de atenção percebida pelo outro olho, fenômeno esse conhecido como confusão (figura 6). Ora, tanto a diplopia do objeto de atenção, quanto à confusão de sua posição espacial com a de outro (espúrio, atrapalhando a visão), são perceptualmente conflitivas, requerendo uma solução. Esta é dada pela supressão das imagens do olho desviado. De fato, tanto a imagem do objeto de atenção, por ele formada na retina periférica, é menos rica em informações discriminativas, quanto a localização espacial de outro objeto, cuja imagem está em sua fóvea, confunde aquela em que se põe o interesse visual; devendo portanto ser, ambas, neutralizadas.

O fenômeno da supressão, aliás, está presente fisiologicamente. Como já comentado, quaisquer objetos fora do horóptero (abrangendo-se aí a área de Panum e a de estereopsia fina) serão percebidos com diplopia (figura 3) e confusão. A atenção concentrada sobre o objeto a ser discriminado, com a supressão natural da duplicação perceptual dos situados além e aquém do horóptero, é o mecanismo pelo qual se evita a queixa de diplopia constante no ato visual corriqueiro e normal. A diplopia na verdade só se torna percebida quando a supressão for neutralizada, o que ocorre quando o objeto de atenção não é visado simultaneamente pelas duas fóveas (estrabismo, incoordenações motoras pelo álcool, etc.).
2) Ambliopia
A continuidade do mecanismo supressivo sobre a percepção das imagens originadas de um só olho (casos de estrabismos monoculares) durante as fases iniciais do desenvolvimento visual, conduz ao estado de ambliopia (do grego, "ambliós", embotado, fraco; "ops", olho, visão). Nela, a principal manifestação clínica é a da redução de acuidade visual, freqüentemente mais declarada no teste feito com os optotipos em tabelas (isto é, com os submetidos às chamadas "interações de contornos") do que com a avaliação por optotipos isolados; propriedade, aliás, considerada patognomônica dessa afecção sensorial.
A importância da ambliopia é ressaltada pelo fato de que as perdas visuais que a acompanham tornam-se irreversíveis se não precocemente tratadas. A recuperação da acuidade visual é provida pelo uso do olho afetado; para que isso ocorra, o método tradicional é o da oclusão do olho bom. Mas como tal procedimento pode levar à ambliopia desse olho, principalmente em tenras idades, são recomendados esquemas em que as oclusões sejam trocadas, com durações para cada um dos olhos que variam em função da idade da criança, da profundidade das perdas visuais e da própria resposta ao tratamento.
A continuidade da supressão, levando à ambliopia, ocorre também nas anisometropias, em que a percepção do ambiente por um dos olhos, com imagens mais desfocadas, deve ser impedida de chegar a níveis de consciência. E também nas aniseiconias em que, mesmo com imagens nítidas, porém de tamanhos desiguais, a fusão ficaria impossibilitada.
Porém, mais graves que as ambliopias causadas pelos mecanismos centrais da supressão (a estrabísmica, a anisometrópica e a aniseicônica) são as que se desenvolvem por ausência de estimulação, ou por sua absoluta inadequação, as de privação ou desuso (ex-anopsia) das quais a causa mais comum é a catarata congênita.
3) Correspondência visual anômala
Outro mecanismo de adaptação sensorial no estrabismo é o do desenvolvimento de um estado em que direções visuais normalmente discrepantes passam a se corresponder. Desse modo, tanto a percepção da diplopia quanto à da confusão tornam-se evitadas.
A correspondência visual (ou retínica) anômala é importante porque pode suscitar diplopia e confusão quando o ângulo (objetivo) de desvio dos eixos visuais (estrabismo) for corrigido por uma cirurgia. Isso se deve a que, nos casos de correspondência visual anômala, ocorre uma dissociação entre a posição dos eixos visuais, cujo ângulo é objetivamente determinado pelo teste de cobertura e prismas (ângulo do estrabismo, desvio oculomotor) e a informação sobre as respectivas percepções (ângulo subjetivo, desvio sensorial). Quando o desvio subjetivo for zero, ou seja, quando o ângulo de anomalia (diferença entre o objetivo e o subjetivo) for igual ao ângulo objetivo, a correspondência visual anômala é dita harmônica. Quando o ângulo subjetivo é diferente de zero (e.g., +15D) e menor que o objetivo (+40D), a correspondência é dita desarmônica (o ângulo de anomalia, nesse caso, é +25D).
A correspondência visual anômala instala-se tanto mais facilmente e é então tanto mais arraigada, quanto menor seja o ângulo do estrabismo. Isso faz sentido, porque é mais fácil o estabelecimento de uma correlação sensorial de elementos de hierarquia mais aproximada (e.g., a fóvea de um olho e um elemento parafoveal do outro) do que entre elementos muito díspares (e.g., a fóvea de um olho e um elemento da retina muito periférica, no outro). Tanto que, para ângulos de dimensões muito reduzidas (microtropias), não estão indicadas as tentativas de correção cirúrgica do estrabismo: a recidiva ao estado anterior de desvio (compensando a diplopia decorrente da tentativa de correção) torna-se regra.
REFERÊNCIAS
1. Howard IP, Rogers BJ. Binocular vision and stereopsis. New York: Oxford University Press; 1995.
2. Duke-Elder S. The eye in evolution. London: Henry Kimpton; 1958.
3. Rosenbaum AL, Santiago AP. Clinical strabismus management. Philadelphia: W. B. Saunders; 1999.
4. Deske-Elder S, Abrams D. Ophthalmic optics and refraction. London: Henry Kimpton; 1970.
5. Tschermak-Seysenegg A. Introduction to physiological optics. Springfield: Charles C. Thomas; 1952.
6. Tyler CW. Sensory processing of binocular disparity. In: Schor CM, Ciuffreda KJ, editors. Vergence eye movements: basic and clinical aspects. Boston: Butterworths; 1983.
7. Loomis JM. Visual space perception: phenomenology and function. Arq Bras Oftalmol 2003;Supl.2:26-9.
8. Palmer DA. Measurement of the horizontal extent of Panum's area by a method of constant stimuli. Opt Acta 1961;8:151-9.
9. Mitchell DE. Retinal disparity and diplopia. Vision Res 1966;6:441-51.
10. Fender D, Julesz B. Extension of Panum's fusional area in binocularly stabilized vision. J Opt Soc Am 1967;57:819-30.
11. Wandell BA. Foundations of Vision. Sunderland: Sinauer Assoc; 1995.
Endereço para correspondência
Faculdade de Medicina de Ribeirão Preto Universidade de São Paulo
Ribeirão Preto (SP) - CEP 14049-900
* Todas as imagens, em olhos emétropes ou amétropes, formam-se na retina. O fato de a construção óptica da imagem do ponto objeto situar-se exatamente sobre ela (nos olhos emétropes ou com a acomodação ajustada) sendo, pois, também puntiforme, ou "fora" dela (aquém, nas miopias; além, nas hipermetropias), sendo então um círculo, é irrelevante para estas considerações.
** Na verdade, os sistemas ópticos possuem dois pontos nodais; o ponto nodal "anterior", ou "primeiro", ou "objeto" e o ponto nodal "posterior", ou "segundo", ou "imagem". No entanto, a distância entre eles ("interstício do sistema" é, no olho, muito pequena (cerca de 0,2 mm) e conseqüentemente negligenciável, dando origem ao conceito de ponto nodal único.
*** Não importa a localização absoluta dessas imagens. Ambas podem estar à direita do observador (como Q, figura 3a), ou à esquerda. Mas o que caracteriza o tipo de diplopia (por exemplo, se homônima ou cruzada) é a disposição relativa das imagens.

How to cite this article:
 ABO is licensed under a Creative Commons Attribution-NonComercial 4.0 Internacional.
ABO is licensed under a Creative Commons Attribution-NonComercial 4.0 Internacional.