Arq. Bras. Oftalmol. 2003; 66 (5): 10.1590/S0004-27492003000600012
Total: 2170
Simon Grondin
DOI: 10.1590/S0004-27492003000600012
ABSTRACT
Visual perception is closely related to and associated with the processing of space. Nevertheless, all events occur at some moment in time, and time needs to be processed in the visual mode. After briefly reviewing the many research avenues in which time and vision are linked, this article focuses on the discrimination of visually marked time intervals, emphasizing work from my own laboratory. It presents a theoretical position, that of a single-clock hypothesis, as well as issues and data that show how visual temporal processing can be approached. These issues are partly related to the fact that time is marked by signals from a single source or by signals provided by distanced sources. In the latter case, different questions arise depending on the signals' exact location. It is known that more distance between flashes can result in longer perceived duration (kappa effect). However, the data reported here, involving signals placed on the same vertical plane, show the opposite effect: briefer perceived duration with more distance between flashes. When flashes are located on the same horizontal plane in different hemi-fields, there is also a lateral effect that may influence time judgments.
Keywords: Time perception; Perceived duration; Time markers; Kappa effect
RESUMO
A percepção visual é relacionada e associada estreitamente ao processamento do espaço. No entanto, todos os eventos ocorrem ao mesmo instante no tempo, e o tempo precisa ser processado no modo visual. Depois de revisar brevemente muitos dos percursos de pesquisa nos quais o tempo e a visão estão ligados, este artigo focou na discriminação de intervalos de tempo demarcados visualmente, enfatizando trabalhos de meu próprio laboratório. Ele apresenta uma posição teórica, a hipótese do relógio único, assim como questões e dados que mostram como o processamento temporal visual pode ser abordado. Estas questões são parcialmente relacionadas ao fato de que o tempo é demarcado por sinais de uma única fonte ou por sinais proporcionados por fontes distantes. No último caso, questões diferentes surgem dependendo da exata localização dos sinais. É conhecido que distâncias maiores entre os flashes podem resultar em maior duração percebido (efeito kappa). Entretanto, os dados relatados aqui, envolvendo sinais localizados no mesmo plano vertical, mostram o efeito oposto: durações percebidas menores com maior distância entre os flashes. Quando os flashes estão localizados no mesmo plano horizontal em diferentes hemicampos, há também um efeito lateral que pode influenciar os julgamentos de tempo.
Descritores: Percepção temporal; Duração percebida; Demarcadores temporais; Efeito kappa
From the electrophysiology of retinal or cortical cells to the Gestalt law of organization of visual stimuli, there are a variety of levels for understanding vision and visual perception. In many cases, the visual phenomena to be understood consist of color, form, depth or movement. As can readily be grasped when considering depth or movement, space is often crucial information for the visual system. However, visual events also occur in sequences, and a number of studies focus on the temporal resolution of these events. After briefly reviewing the fundamental temporal properties of the visual system, this article examines the processing of time as information within the visual system.
TIME-RELATED PROPERTIES OF VISUAL PERCEPTION
This section describes various basic time-related issues associated with the visual system. For the sake of simplicity, it is divided into two parts, according to whether or not space is central to the description of the phenomenon reported.
Visual time without space
Many fundamental phenomena in the visual system are determined by the duration of signals or are related to time, given that they continue for a while after a signal's physical offset. Indeed, as noted by Dixon and Di Lollo(1), the visual system must reconcile two opposing, basic requirements, namely, the maintenance of continuity of perception over time (integration) and the detection of rapid physical changes (segregation of stimuli in a sequence).
The perceived duration of signals and the closely related phenomenon of temporal persistence are among the most frequently studied time-related properties of the visual system. The former refers to the period from the stimulus onset to the end of the sensation, while the latter is defined as the period from the signal offset to the termination of the sensation(2). Some authors emphasize the distinction between visible persistence, i.e. the perceptual experience that looks like the continuity of a stimulation beyond the presentation of a brief visual stimulus, and informational persistence, which is the information that can be acquired for the stimulus for a short time after the onset of the physical stimulus(3).
Some very robust phenomena have been reported about visible persistence. One is called the inverse-duration effect,given that the persistence is inversely related to the duration of the stimulus(4). Another phenomenon, more prone to inconsistent data, is the inverse-intensity effect, which refers to the fact that an increase in the intensity of a stimulus may result in reduced visible persistence. This effect is reviewed in depth by Nisly and Wasserman(2). Di Lollo and Bischof(4) also provide a detailed review of the phenomenon, emphasizing the distinction between experiments involving stimuli of varying intensity presented on a fixed background and those involving stimuli of fixed intensity presented on a varied background. Di Lollo and Bischof(4) also argue that increasing intensity may sometimes improve performance, but not visible persistence.
A related inverse-intensity effect is called the critical frequency at fusion, i.e. the ability to distinguish a light that is flickering between on and off states(4). Depending on several conditions such as retinal location, stimulus size or luminance, it is possible to detect flickering even when a light is turned on and off every 10 to 50 ms.
In addition to discussing these visual persistence phenomena, the literature also focuses on the fact that perceptual information is processed in moments that are temporally discrete(5). This concept is referred to as the perceptual moment. According to Stroud, this moment lasts about 100 ms, which would be the minimum perceived duration a stimulus could have: within this range two stimuli would be perceived as simultaneous. Since Stroud published his results, several estimates of the value of a perceptual moment have appeared in the literature. The concept of a perceptual moment is interesting because there are certainly limitations in time regarding the processing of rapidly presented stimuli. However, quantifying the value of this moment precisely seems to be an impossible task given that it fluctuates with the experimental conditions under investigation.
Time embedded in space
The temporal characteristics of signals that reach the eyes and the perception of time in the visual mode are some of the fundamental questions discussed in the present article. However, as soon as the question of time is addressed, it is difficult to avoid relating it to that of space, be it in the realm of physics(6) or from an phenomenological viewpoint(7). In psychology, time and space issues take several forms.
In the visual system, when signals are delivered at several locations in space, some variation in their duration and intensity and in the time between them can create an impression of movement. In other words, just by flashing separate objects on and off, it is possible to produce the impression that something is moving even when no object is moving at all. Examples of this phenomenon were reported almost a century ago by Korte and Wertheimer (see also, for instance, Burt and Sperling(8)). This impression of movement is generally called apparent movement, and sometimes stroboscobic movement, beta movement or the phi phenomenon.
A totally different approach to the space/time issue has emerged in the literature on time-to-collision (also called time-to-contact). With this phenomenon, there is real movement, i.e. the displacement of an object in space. The success of a time-to-collision task requires some implicit timing. For instance, catching a ball requires certain spatial and temporal calculations of the displacement of both the observer and the ball(9-11). Several other sports activities, such as diving or batting, require temporal adjustments to a point in space. As well, stopping a car safely at a given point requires the ability to correctly distribute the force applied to the brakes over time and, therefore, some perspective of time(12). The literature provides several theoretical analyses of the variation of optical information prior to an upcoming contact(13-14).
TIME AS INFORMATION
The key issue in this section is the processing of time. In other words, time is treated like explicit information that has to be processed. Empirical facts about duration discrimination in the visual mode will be reviewed, according to whether one or more sources of light provide the signals for marking the intervals to be processed.
Timing mechanisms
Before reviewing empirical facts about the discrimination of visually marked intervals, this article will briefly discuss the main contemporary theories about temporal processing.
There are many models for explaining how time judgments are made. Some recent proposals are based on oscillatory processes(15-18), while others emphasize the temporal regularity of events in the environment(19) or the role of memory(20).
The most popular contemporary models of time perception are probably those based on a clock-counter device(21-23) (for a review, see Grondin(24)). This class of models is embedded in the general information processing version of Scalar Timing Theory(25), developed mainly by Gibbon, Church and Meck(26) (see also Treisman(27)). Within this framework, the variability of temporal judgments is located at three processing levels, as indicated by figure 1.
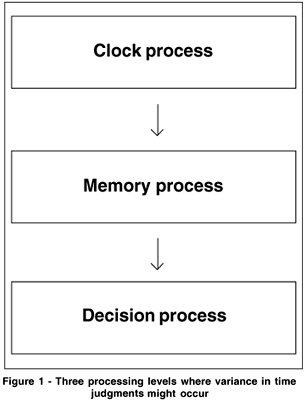
In some parts of the empirical work presented below, the first component, or the clock process of this general framework, is the most interesting aspect. The clock itself is usually described as a pacemaker-counter device (Figure 2). Its functioning is based on the emission of pulses of some sort by a pacemaker, and the number of pulses accumulated by the counter determines the perceived duration of a temporal interval. The pacemaker-counter device is frequently reported to include a switch component that can interrupt the transmission of pulses to the counter. This switch component is often said to be under control of attention(28) (for a review(29)).

Many fundamental questions emerge from this clock concept. One relates to the clock's pulse emission properties which, depending on the model in question, are considered to be deterministic or stochastic. In the latter case, the clock is sometimes assumed to be a Poisson process(21,30-31). While it is often assumed that the counter process is error free, some authors report that this component involves variance(32-33).
Temporal discrimination with markers from a single source
Many experiments use only one source of light for marking time. Intervals can be defined by the period a flash is lit (filled interval) or by the duration between two brief flashes, from the onset or offset of the first signal to the onset of the second signal (empty interval).
The discrimination of very brief intervals, either filled or empty, has been under investigation. With empty intervals marked by 1-ms pulses, Nilsson(34) showed that, contrary to Weber's law,1 the threshold did not rise with increases of up to 30 ms in base intervals. Moreover, his study revealed that certain aspects of visual stimuli interfere with the processing of time.
In contrast, Allan, Kristofferson and Wiens(35) used filled visual intervals for studying brief interval discrimination. Their experiments are important because they showed that participants, in a range of duration where Bloch's law is known to apply, can really base their judgments on the temporal extent of visual signals, and not on the signals' intensity. Bloch's law (or the Bunsen-Roscoe law) states that, for very brief visual stimuli (less than.1 s), the detection threshold depends on the reciprocal relation between the duration and intensity of signals.
When longer intervals are studied, duration discrimination in the visual mode follows the Generalized form of Weber's law, with either filled or empty intervals(36), with a clear constancy of the Weber fraction from 500 to 740 ms (with empty intervals(37)) and probably up to 900 ms(38). However, above 900 ms, there is a discontinuity of the function if participants are asked to refrain from using explicit counting during the discrimination task and if intervals are marked by sequences of brief flashes. In other words, the Weber fraction increases with longer intervals(38).
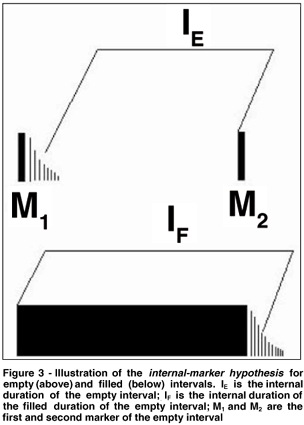
Moreover, for intervals ranging from 250 to 500 ms, temporal discrimination is usually better with empty intervals than with filled ones(36); the difference disappears, however, with longer intervals(39). The filled/empty difference can probably be explained by what has been called an internal-marker hypothesis(36,40). According to this hypothesis, discrimination levels for filled and empty intervals may differ because the timekeeping period in each case is not equal.
Even if the physical duration of filled and empty intervals is identical, the ways of marking time internally might not be the same. The difference may be related to the signals' response characteristics. On the one hand, the beginning of the internal timekeeping period might be about the same if the observer starts timing at the onset of a signal. Indeed, with a filled interval, timekeeping starts at the detection of the visual signal, while, with an empty interval, the timekeeping activity might only start when the signal fades. On the other hand, with empty intervals, the timekeeping activity stops when the second signal occurs. This is not the case with filled intervals, where the observer stops counting time when the visual signal disappears (see above-mentioned concept of visible persistence). In other words, performance differences may result from (1) the influence of different non-clock sources of variance on the timing mechanism and (2) the fact that the timekeeping period differs for filled and empty intervals. In the first case, the variability of sensory signals, marking the beginning or end of the internal timekeeping period, contributes to the overall variance of the process. In the second case, it is the clock-based variance that differs with the interval structure since the timekeeping period differs: a longer time keeping period results in higher temporal variance. As shown in figure 3, a filled interval might actually be perceived as being longer than an empty one, which is consistent with the timing literature(41-42).
Finally, it is important to stress that major distortion of perceived duration can be caused by the presentation of successive visual signals. For instance, when a series of 600-msec light flashes were used to mark filled visual intervals, the first of a series of four intervals was overestimated by as much as 50%; i.e. it would have to be shortened considerably to appear to have the same duration as the following intervals(43). This distortion, which is reported to be specific to the visual system, applies to a variety of stimulus contexts and sizes and inter-flash interval conditions. Arao, Suetomi and Nakajima(44), for their part, have reported specific conditions where substantial underestimation of empty intervals can occur: when two time intervals are marked by three successive brief flashes, the second interval is perceived as being much shorter because of the presence of the preceding interval. This phenomenon, called time-shrinking, has already been well-documented for the auditory system(45-46). In the visual system, it usually occurs when the second interval is longer than the first one and the first interval ranges from 160 to 400 ms.
Temporal discrimination with markers from multiple sources
Although the sensory effects reported in the previous section are still potential sources of variance when more than one visual source is used for marking temporal intervals, the main concern in this section is that a fundamental phenomenon, namely, space, may interfere with the timing process when two or more light sources are involved in a visual discrimination task. The potential influences of space on time judgments are reviewed below.
A classical case: kappa
Different streams of literature on visual perception in experimental psychology, which focus, for example, on the above-mentioned concepts of time-to-collision or apparent movement, include a variety of time and space considerations. One such consideration, namely, the kappa effect, is of special interest as it is directly related to explicit judgments about time(47). Time judgments are influenced by the distance between visual sources marking time. The mirror image of this effect, called the tau effect(48), occurs when duration affects perceived distance. The kappa effect is reported to exist not only with visual signals, but also with tactile ones(48), and also in some form in the auditory system(50).
The kappa effect is usually shown to be robust in conditions where three successive signals (such as X, Y and Z, with Y somewhere between X and Z) are delivered. For two equal time intervals defined by the onset of two signals, X-Y or Y-Z, duration is perceived as longer for the X-Y sequence than for the Y-Z one if the distance between X and Y is greater than that between Y and Z. However, this kappa effect seems to appear only under certain experimental conditions. It may not apply to very brief durations, i.e. when the addition of X-Y and Y-Z equals 160 ms(51-53), and it is more likely to occur under difficult discriminability conditions(53).
The vertical case: depth effect?
The kappa effect is usually present with a sequence of three flashes marking two intervals. In this context, more space between signals is shown to lead to longer perceived duration. In a recent experiment(54), this distance effect was investigated with a different procedure. Three signals located on the same vertical plane were used, with one flash placed midway between the other two. If space is taken into account as in the kappa effect, a sequence marked by the upper and lower signals should be perceived as being longer than signals involving the mid-point flash. Furthermore, given that what is higher in the visual field is usually perceived as being farther away(55), it was expected that intervals marked by the middle and upper flashes would be perceived as longer than those marked by the middle and lower flashes.
In the experiment, twenty-four participants had to categorize temporal intervals as short or long. The intervals were marked by two brief visual signals. The signals, LEDs lit for 20 ms, were delivered from three potential locations on a vertical plane in front of the participants, i.e. above (A), middle (M) and below (B), with about 25 cm between A and M, and between M and B. The visual stimuli were placed about 1 m in front of the participants, subtending a visual angle of about. 57°. The eye level of participants was at the height of M, and participants were asked to look at M.
The categories assigned to intervals marked by M-A, B-M and B-A sequences (ascending trials) were compared, as were those assigned to intervals marked by A-M, M-B and A-B sequences (descending trials). Two ranges of duration, 160 and 320 ms, and two critical dependent variables, the constant error (CE) and the standard deviation (SD)2, were studied.
Mean CE and SD are reported in Figure 4 for the ascending trials (upper panel) and for the descending ones (lower panel). In both cases, there is a significant signal location effect for CE: more distance between signals results in shorter perceived duration. For SD, the results show significant location effects only in the descending trials: discrimination was better in the M-B condition than in the A-M one.
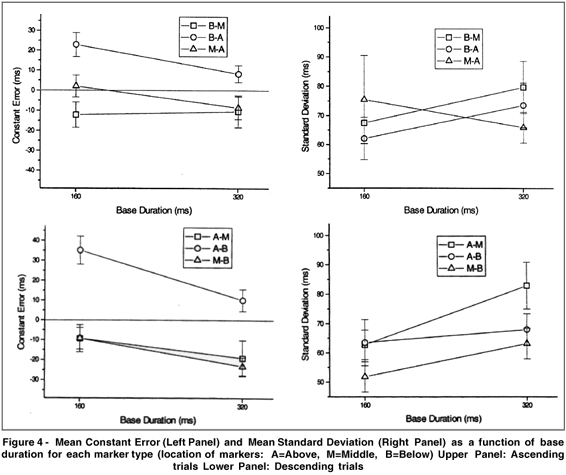
The greater the space between signals (A-B or B-A), the shorter the perceived duration. This finding is not consistent with the kappa effect described earlier. On the contrary, in both ascending and descending trials, the results demonstrate that duration was perceived as being shorter when signals were at a greater distance from one another. Under these circumstances, it is not surprising that none of the results support the second prediction, that is, the one regarding the relative height of signals, i.e. the A-M vs. M-B or M-A vs. B-M comparison. In short, the main factor in the experiment was not the relative height of signals, but the total distance between them.
This distance effect can be explained by the internal-clock perspective described above. It is often assumed that one of the critical factors determining the accumulation of pulses in an internal-clock system is whether or not attention is paid to time(57-58). In this context, the present CE results might be interpreted as indicating that a greater distance condition would require a larger displacement of attention from one signal to another. This switching process would detract attention from time, diminishing the number of pulses accumulated during the interval to be timed.
Phenomena such as apparent movement or the kappa effect demonstrate the non-independence of space and time. Kappa in particular reveals that more space between events results in longer perceived duration. However, some conditions such as those described above clearly show that more space between events can also result in an impression that time is briefer. Future research is needed to provide a clearer picture about the conditions under which space and, especially, distance or depth cues influence the perception of time between the occurrence of successive visual signals.
The horizontal case: laterality effect?
When different sources of light marking time are located on a horizontal plane, an additional theoretical issue may emerge: if these sources are placed in different visual hemi-fields, the flashes are processed by different cerebral hemispheres. This type of experimental setting leads to the question of the lateralization of temporal processing mechanisms. The literature on the effects of laterality on temporal processing provides some evidence of the superiority of the left cerebral hemisphere over the right(59). Nicholls(60) recently conducted an extensive review of the literature on the temporal processing asymmetries that occur between cerebral hemispheres. In this review, most of the evidence for the superiority of the left hemisphere (LH) for processing time derives from the results obtained in tasks such as: 1) judging the temporal order of two stimuli; 2) judging the simultaneity, as opposed to the succes siveness, of two stimuli; and 3) detecting a gap in a signal. In general, the left cerebral hemisphere is reported to be superior in such tasks, when signals are delivered in the visual, auditory or tactile modality.
The above-mentioned tasks involve time-related processing, but not necessarily the processing of time itself. There is no evidence in support of left hemisphere superiority in terms of duration discrimination. The few data available on this issue suggest no cerebral difference in the performance of this task when visual markers are used(61), and even some neuropsychological evidence of a superiority of the right hemisphere for auditory temporal discrimination(62-63).
In Grondin(61), two visual signals (left or right) established four marker-type empty intervals, two of which were presented unilaterally (left-left or right-right) and the other two, bilaterally (left-right or right-left). It was shown that perceived duration is longer with a left-right sequence than with a right-left sequence. This is not compatible with the left-hemisphere hypothesis for temporal processing, but it does comply with the hypothesis of an internal mechanism that scans visual information in a left-right order(64). In Grondin(61), it was also shown that discrimination is better with unilateral sequences than with bilateral ones.
More recently, an experiment compared discrimination levels in conditions where intervals (either filled or empty) were presented unilaterally (with two LEDs) or marked by one LED in each hemi-field at the same time(65). Using signals in both hemi-fields was expected to favor the contribution of both hemispheres to the time-processing task, a phenomenon that might enhance performance. A somewhat related finding was reported by Helmuth and Ivry(66) for the production of intervals in a repetitive timing task. However, depending on its structure (filled vs. empty), an interval may require different types of processing, i.e. analytic or holistic, which are reported to be the domain of the left and right cerebral hemispheres respectively(67). Participants in Grondin, Guay et al.(65) were asked to discriminate between 50- and 80-, 100- and 150-, or 210- and 290-ms intervals.
As indicated in Figure 5, which illustrates the mean percentage of correct responses in each experimental condition, the results tend, in all conditions, toward better discrimination when signals are delivered from the RVF than from the LVF or from both VFs. However, none of the results are significant. Moreover, the superiority of one interval structure over the other (filled vs. empty) depends on the range of duration under study. This structure effect is significant only with briefer intervals.
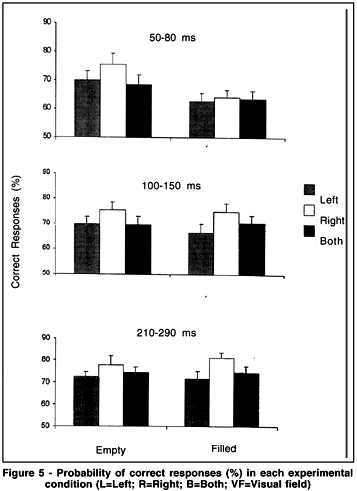
At 50-80 ms, having better discrimination with empty intervals than with filled ones is consistent with Grondin(36), where a quite similar method was employed; however, the level of performance in the present experiment was much lower. This might be ascribed to the fact that the signals were presented at periphery in the present study, but at fovea (only one LED signal) in Grondin(36). In other words, the overall level of temporal performance is not independent of the quality of a sensory impression (fovea vs. periphery). Indeed, overall discrimination is better for 250-ms empty intervals when both visual markers are presented at fovea than it is in the periphery conditions(61).
Future research on lateral effects in visual temporal processing should focus on rhythmic effects, i.e. on the impact of using sequences of flashes. Ben-Dov and Carmon(68) used a task where two separate sequences of intervals marked by brief flashes had to be categorized as being the same or different. A short (S=.2 s) and a long (L=.4 s) interval were presented. For instance, a SLS sequence was followed by a SLS sequence (same) or a SSL sequence (different). The main dependent variable used was reaction time. Ben-Dov and Carmon(68) showed that reaction time was faster for signals presented in the right visual field (left-hemisphere –LH— superiority) when the sequence contained fewer elements, but faster for signals presented in the left visual field (right-hemisphere - RH - superiority) when the sequence contained more elements, i.e. when a rhythm was induced. This finding was attributed to the distinct modes of information processing, that is, analytic and holistic, used by the LH and RH respectively.
CONCLUDING REMARKS
A more complete understanding of information processing in the visual system will probably be achieved when certain temporal features of visual signals are better understood. These features include the time needed to integrate or segregate visual signals and the impact of these signals on explicit judgments about time. One of the critical issues relating to explicit temporal judgments is the integration of spatial parameters. Depending on the spatial location of visual signals marking time, different perspectives must be used for interpreting data. First, the exact conditions where the kappa effect occurs or does not occur should be more clearly determined. In addition, any theoretical explanation of how space exerts an effect on perceived time must take into account: (1) the availability of depth cues and the magnitude of these cues' effect and (2) the possibility of cerebral hemispheric superiority or specialization in the processing of time.
AUTHOR'S NOTE
The research was supported by a Grant from the Natural Sciences and Engineering Council of Canada
REFERENCES
1. Dixon P, Di Lollo V. Beyond visible persistence : An alternative account of temporal integration and segregation in visual processing. Cognit Psychol 1994;26:33-63.
2. Nisly TH, Wasserman GS. Intensity dependence of perceived duration : Data, theories, and neural integration. Psychol Bull 1989;106:483-96.
3. Loftus GR, Irwin DE. On the relations among different measures of visible and informational persistence. Cognit Psychol 1998;35:135-99.
4. Di Lollo V, Bischof WF. Inverse-intensity effect in duration of visible persistence. Psychol Bull 1995;118:223-37.
5. Stroud JM. The fine structure of psychological time. In: Quastler H, editor, Information theory in psychology: Problems and methods. Glencoe: Free Press; 1955, p.174-205.
6. Buccheri R, Saniga M. The nature of time: Geometry, physics, and perception. Dordrecht: Kluwer, in press.
7. Delong AJ. Phenomenological space-time: Toward an experiential relativity. Science 1981;213:681-2.
8. Burt P, Sperling G. Time, distance, and feature trade-offs in visual apparent motion. Psychol Rev 1981;88:171-95.
9. Oudejans RRD, Michaels CF, Bakker FC, Davids K. Shedding some light on catching in the dark: Perceptual mechanisms for catching fly balls. J Exp Psychol Hum Percept Perform 1999;25:531-42.
10. Sheffer DM, McBeath MK. Baseball outfielders maintain a linear optical trajectory when tracking uncatchable fly balls. J Exp Psychol Hum Percept Perform 2002;28:335-48.
11. Tresilian JR. Perceptual and cognitive processes in time-to-contact estimation: Analysis of prediction-motion and relative judgment task. Percept Psychophys 1995;57:231-45.
12. Lee DN. A theory of visual control of braking based on information about time-to-collision. Perception 1976;5:437-59.
13. Kaiser MK, Phatak AV. Things that go bump in the light: On the optical specification of contact severity. J Exp Psychol Hum Percept Perform 1993; 19:194-202.
14. Kim N-G, Turvey MT, Carello C. Optical information about the severity of upcoming contacts. J Exp Psychol Hum Percept Perform 1993;19:179-93.
15. Buonomano DV, Merzenich MM. Temporal information transformed into a spatial code by a neural network with realistic properties. Science 1995;267: 1028-30.
16. Large E, Jones M. The dynamics of attending: How people track time-varying events. Psychol Rev 1999;106:119-59.
17. Miall C. Models of neural counting. In: Pastor MA, Artieda J, editors, Time, internal clocks and movement. Amsterdam: Elsevier; 1996, p.69-94.
18. Treisman M, Faulkner A, Naish PLN, Brogan D. The internal clock: Evidence for a temporal oscillator underlying time perception with some estimates of its characteristic frequency. Perception 1990;19:705-43.
19. Jones MR, Boltz MG. Dynamic attending and responses to time. Psychol Rev 1989;96:459-91.
20. Staddon JER, Higa JJ. Time and memory: Towards a pacemaker-free theory of interval timing. J Exp Anal Behav 1999;71:215-51.
21. Gibbon J. Origins of scalar timing. Learn Motiv 1991;22:3-38.
22. Killeen PR, Weiss NA. Optimal timing and the Weber function. Psychol Rev 1987;94:455-68.
23. Rammsayer TH, Ulrich R. Counting models of temporal discrimination. Psychon Bull Rev 2001;8:270-7.
24. Grondin S. From physical time to the first and second moments of psychological time. Psychol Bull 2001a;127(1):22-44.
25. Allan LG, Church RM. In memoriam: John Gibbon (1934-2001). Learn Motiv (Special issue) 2002;33:1-8.
26. Gibbon J, Church RM. Sources of variance in an information processing theory of of timing. In: Roitblat HL, Bever TG, Terrace HS, editors, Animal cognition. Hillsdale: Erlbaum; 1984, p.465-88.
27. Treisman M. Temporal discrimination and the indifference interval: Implications for a model of the "internal clock". Psychol Monographs 1963;77: Whole 576.
28. Meck WH. Attentional bias between modalities: Effect on the internal clock, memory, and decision stages used in animal time discrimination. In: Gibbon J, Allan LG, editors, Annals of the New York Academy of Sciences, vol. 423 – Timing and time perception. New York: New York Academy of Sciences; 1984, p.528-41.
29. Grondin S. Time psychophysics and attention. Psychologica 2001b;28:177-91.
30. Creelman CD. Human discrimination of auditory duration. J Acoust Soc Am 1962;34:582-93.
31. Divenyi PL, Danner WF. Discrimination of time intervals marked by brief acoustic pulses of various intensities and spectra. Percept Psychophys 1977; 21:125-42.
32. Killeen PR, Taylor T. How the propagation of error through stochastic counters affects time discrimination and other psychophysical judgments. Psychol Rev 2000;107:430-59.
33. Killeen PR, Fetterman JG. A behavioral theory of timing. Psychol Rev 1988; 95:274-95.
34. Nilsson TH. Two-pulse interval vision thresholds. J Opt Soc Am 1969;59: 753-6.
35. Allan LG, Kristofferson AB, Wiens EW. Duration discrimination of brief light flashes. Percept Psychophys 1971;9:327-34.
36. Grondin S. Duration discrimination of empty and filled intervals marked by auditory and visual signals. Percept Psychophys 1993;54:383-94.
37. Grondin S, Ouellette B, Roussel M-È. About optimal timing and stability of Weber fraction for duration discrimination. Acoust Sci Technol 2001;22:370-2.
38. Grondin S. Discriminating time intervals presented in sequences marked by visual signals. Percept Psychophys 2001c;63:1214-28.
39. Grondin S, Meilleur-Wells G, Ouellette C, Macar F. Sensory effects on judgments of short-time intervals. Psychol Res 1998;61:261-8.
40. Grondin S, Ivry RB, Franz E, Perreault L, Metthé L. Markers' influence on the duration discrimination of intermodal intervals. Percept Psychophys 1996; 58:424-33.
41. Goldfarb JL, Goldstone S. Time judgment: A comparison of filled and unfilled durations. Percept Mot Skills 1963;16:376.
42. Goldstone S, Goldfarb JL. Judgment of filled and unfilled durations: Intersensory factors. Percept Mot Skills 1963;17:763-74.
43. Rose D, Summers J. Duration illusions in a train of visual stimuli. Perception 1995;24:1177-87.
44. Arao H, Suetomi D, Nakajima Y. Does time-shrinking take place in visual temporal patterns? Perception 2000;29:819-30.
45. Nakajima Y, ten Hoopen G, van der Wilk RGH. A new illusion of time perception. Music Percept 1991;8:431-48.
46. ten Hoopen G, Hilkhuysen G, Vis G, Nakajima Y, Yamauchi F, Sasaki T. A new illusion of time perception – II. Music Percept 1993;11:15-38.
47. Jones B, Huang YL. Space-time dependencies in psychophysical judgment of extent and duration: Algebraic models of tau and kappa effects. Psychol Bull 1982;91:128-42.
48. Helson H. The tau effect: An example of psychological relativity. Science 1930;71:536-7.
49. Suto Y. The effect of space on time estimation (S effect) in tactual space. II. The role of vision in the S effect upon skin. Jpn J Psychol 1955;26:94-9.
50. Cohen J, Hansel CEM, Sylvester JD. A new phenomenon in time judgment. Nature 1953;172:901.
51. Collyer CE. Discrimination of spatial and temporal intervals defined by three light flashes: Effects of spacing on temporal judgments and of timing on spatial judgments. Percept Psychophys 1977;21:357-64.
52. Huang YL, Jones B. On the interdependence of temporal and spatial judgments. Percept Psychophys 1982;32:7-14.
53. Myatani M. The time and distance judgments at different levels of discriminability of temporal and spatial information. Hiroshima Forum Psychol 1985; 10:45-55.
54. Guay I, Grondin S. Influence of time interval categorization of distance between markers located on a vertical plane. In: Sommerfeld E, Kompass R, Lachmann T, editors, Proceedings of the 17th Annual Meeting of the International Society for Psychophysics. Berlin: Pabst Science Publishers; 2001, p.391-6.
55. Girgus JS, Coren S. Depth cues and constancy scaling in the horizontal-vertical illusion: The bisection error. Can J Psychol 1975;29:59-65.
56. Ivry RB, Hazeltine RE. The perception and production of temporal intervals across a range of durations: Evidence for a common timing mechanism. J Exp Psychol Hum Percept Perform 1995;21:3-18.
57. Grondin S, Macar F. Dividing attention between temporal and nontemporal tasks: a performance operating characteristic – POC – analysis. In: Macar F, Pouthas V, Friedman W, editors, Time, action, cognition: towards bridging the gap. Dordrecht: Kluwer; 1992, p.119-28.
58. Macar F, Grondin S, Casini L. Controlled attention sharing time estimation. Mem Cognit 1994;22:673-86.
59. Efron R. The decline and fall of hemispheric specialization. Hillsdale: Erlbaum, 1990.
60. Nicholls MER. Temporal processing asymmetries between the cerebral hemispheres: evidence and Implications. Laterality 1996;1:97-137.
61. Grondin S. Judgments of the duration of visually marked empty time intervals: Linking perceived duration and sensitivity. Percept Psychophys 1998;60:319-30.
62. Harrington DL, Haaland KY, Knight RT. Cortical networks underlying mechanisms of time perception. J Neurosci 1998;18:1085-95.
63. Milner B. Laterality effects in audition. In: Mountcastle VR, editor, Interhemispheric relations and cerebral dominance. Baltimore: John Hopkins University Press; 1962, p.177-95.
64. Sekuler R, Tynan P, Levinson E. Visual temporal order: A new illusion. Science 1973;180:210-2.
65. Grondin S, Guay I, Lapointe M, Ouellet B. About Hemispheric Asymmetries in Relation to the Discrimination of Visually Marked Intervals. Brain Cogn 2002;48:(abstract).
66. Helmuth LL, Ivry RB. When two hands are better than one: Reduced timing variability during bimanual movements. J Exp Psychol Hum Percept Perform 1996;22:278-93.
67. Bradshaw JL, Nettleton NC. The nature of hemispheric specialization in man. Behav Brain Sci 1981;4:51-91.
68. Ben-Dov G, Carmon A. Rhythm length and hemispheric asymmetry. Brain Cogn 1984;3:35-41.
Correspondence to
Simon Grondin
École de Psychologie, Université Laval, Québec
Québec, Canada, G1K 7P4
email:[email protected]
phone: (418) 656-2131 ext. 6227 / Fax: (418) 656-3646
1 According to Weber's law, the minimum difference, · f, in magnitude required between two stimuli in order to distinguish them increases monotonically as a function of the magnitude of a standard duration, f The ratio, · f/f, is called the Weber fraction and should be constant according to Weber's law. Essentially, the Generalized form of Weber's law states that this ratio could be higher for low values of f.
2 In this experiment, where marking signals were located in a vertical plane, a 6-point psychometric function was traced for each subject and each experimental condition, plotting the six comparison durations used for one standard condition on the x axis and the probability of responding "long" on the y axis. The cumulative normal distribution was fitted to the resulting curves. The bisection point, BP, i.e. the point on the x axis where the probability of responding "long" is.50, was estimated for each experimental condition. The BP minus the base duration (160 or 320 ms) gives the constant error (CE). Note that a higher CE value indicates a shorter perceived duration (more "short" responses). Also, one SD on the psychometric function indicates sensitivity for categorizing intervals as short or long. One (1) standard deviation (SD) is commonly used to express sensitivity in time research(22,38,55).

How to cite this article:
 ABO is licensed under a Creative Commons Attribution-NonComercial 4.0 Internacional.
ABO is licensed under a Creative Commons Attribution-NonComercial 4.0 Internacional.